

Génétique / Breeding
By Cr4b
Sommaire :
1. Introduction à la génétique
1.1. Notion de gène
1.2. Notion d’allèles
1.3. Notion de méiose
1.4. Notion de lignées pures
1.5. Notion de Génotype/Phénotype
Notations usuelles
2.1. Effet de la dominance dans un croisement mono hybride
2.2. Exemple d’un rétro croisement F1 et P1
3. Dihybridisme/Polyhybridisme
. Transmission de deux paires de gènes indépendants
4. Les limites de la génétique mendélienne
4.1. Dominance
4.2. Interaction génique
4.3. Transmission indépendante
5. Applications à la reproduction sélective
7. Hybridation et Différenciation
8.1. Le goût d'ananas est dominant
8.2. Le goût d'ananas est récessif
8.3. Appliquer des critères de sélection
8.4. Cubing sélectif
8.5. En conclusion
9.1. Stabilisation à partir d'un seul individu
9.2. La stabilisation par croisements successifs
. Liste des caractéristiques favorables observées chez le cannabis et sujettes à variations
12.1. Sélection des parents
12.2. Récolte du pollen
12.3. Conservation du pollen
12.4. Pollinisation
13.1. Maturité des graines
13.2. Sélection des graines
13.3. La levée de dormance
14.1. De la théorie à la pratique
14.2. Sur combien de pieds travailler ?
15. Le cas particulier de la polyploïdie
15.1. Conséquence de la polyploïdie
15.2. Obtenir des plants diploïdes [conseil]
1 - Introduction à la génétique
Le cannabis est une plante généralement dioïque de nombre haploïde (1n) de chromosomes égal à 10 et de nombre diploïde (2n) égal à 20 (Mendel, 1964)
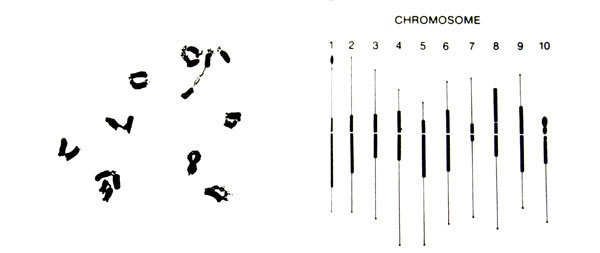
1.1 - Notion de gène
Un gène désigne une unité d'information génétique transmise par un individu à sa descendance, par reproduction sexuée ou asexuée. Le gène le plus simple consiste en un segment d'acide nucléique codant pour une seule protéine ou un ARN (en dehors de l'épissage alternatif). L'ensemble des gènes d'un individu constitue son génome. Les gènes ne constituent qu'une partie du génome.
Plus généralement, le terme est utilisé relativement à la transmission et à l'hérédité de caractères identifiables particuliers.
Un gène est donc une unité d'information génétique qui permet la synthèse d'un polypeptide.
Un gène est caractérisé par sa séquence de nucléotides et le polypeptide par sa séquence en acides aminés, on peut penser que la séquence de nucléotides du gène doit déterminer la séquence d'acides aminés du polypeptide pour lequel il code.
1.2 - Notion d’allèles
On nomme allèle une variante donnée d'un gène au sein d'une espèce.
Dans une cellule diploïde, il y a deux allèles pour chaque gène : un allèle transmis par chaque parent.
Les allèles transmis par les parents peuvent être identiques ou non.
Si les allèles apportés par chaque parent sont identiques dans leur séquence nucléotidique, l'individu est homozygote pour ce gène.
S’ils sont différents, l'individu est hétérozygote.
Dans ce dernier cas, deux possibilités sont envisageables quant au phénotype résultant de l'expression du gène.
Si l'un des deux allèles s'exprime et l'autre reste « muet », on dit que le premier est dominant et l'autre récessif. Les allèles dominants sont symbolisés par une lettre majuscule, et les récessifs par une lettre minuscule.
1.3 - Notion de méiose
La méiose est un processus se déroulant durant la gamètogenèse (spermatogenèse ou ovogenèse), c'est-à-dire durant l'élaboration des gamètes. Elle a pour but de donner des cellules haploïdes à partir de cellules diploïdes au cours de deux divisions.
Mais en plus de ce rôle de division, la méiose a un rôle important dans le brassage génétique et ce à cause de deux brassages : Le brassage interchromosomique et le brassage intrachromosomique.
Ainsi, durant la méiose, la quantité d'ADN au sein de la cellule évolue au cours du temps. Nous pouvons aisément le voir à l'aide d'un graphique :
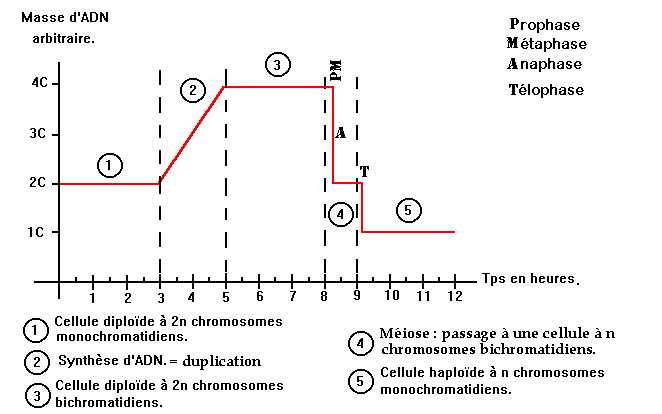
1.4 - Notion de lignées pures
Une lignée est dite « pure » si le croisement des deux parents possédant un caractère commun déterminé génétiquement (ou l’auto-pollinisation d’une plante monoïque) donne une descendance présentant le même caractère et si toutes les générations suivantes « endogames » le présentent à leur tour.
On en déduit que les parents sont homozygotes et qu’ils possèdent le même allèle.
1.5 - Notion de Génotype/Phénotype
Le génotype est le patrimoine héréditaire d'un individu. Tous les individus d’une même espèce possèdent le même nombre de gènes (on estime environ 50.000 à 100.000 le nombre de gènes chez l’espèce humaine).
Pour chacun de ces gènes, il existe des versions différentes appelées allèles. Les gènes sont portés par les chromosomes (23 paires chez l'espèce humaine; notation : 2N=46) transmis par les cellules sexuelles.
On appelle génotype l’ensemble des allèles d’un individu portés par l'ADN d'une cellule vivante.
L'interaction du génotype d'un individu avec son environnement détermine son phénotype. C'est donc une information qui intervient au niveau des gènes et qui peut-être modifiée par une mutation.
Notations usuelles
On va désigner par :
- une lettre majuscule les allèles dominants
- une lettre minuscule les allèles récessifs
- P, les parents
- F, les lignées filles
- S, les graines issues d'un selfing
- BC, les graines issues d'un back cross
2 - Monohybridisme
On va étudier le cas d’un croisement entre deux parents issus de variétés pures différentes pour un caractère donné.
Dans ce cas, les descendants seront des hybrides dont les variations peuvent être prédites par la génétique mendélienne.
Prenons l’exemple de deux lignées de cannabis pures pour la caractère forme des feuilles.
La lignée 1 est WW avec W, allèle dominant responsable de feuille penniforme
La lignée 2 est ww, avec w allèle récessif responsable de feuille palmée.
2.1 - Effet de la dominance dans un croisement mono hybride

Par contre, il s’agit ici d’un cadre très restrictif où la dominance est totale et la forme des feuilles contrôlée uniquement par un seul gène.
Cela a aussi un intérêt dans la détermination du génotype parental vu que nous n’avons accès qu’au phénotype.
2.2 - Exemple d’un rétro croisement F1 et P1

3 - Dihybridisme/Polyhybridisme
Ces règles peuvent s’appliquer à plusieurs gènes, dans ce cas on va parler de polyhybridisme.
Prenons l’exemple de deux gènes se transmettant de façons indépendantes. Avec :
- T allèle dominant codant pour un phénotype grand
- t, allèle récessif codant pour un phénotype court
- M, allèle dominant codant pour un phénotype précoce
- m , allèle récessif codant pour un phénotype tardif
Transmission de deux paires de gènes indépendants :

On s’aperçoit qu’en F2 apparaissent deux phénotypes non parentaux (grand/tardif et court/précoce). Il s’agit du phénomène dit de recombinaison.
4 - Les limites de la génétique mendélienne
Il s’agit ici du cadre très restrictif de la génétique mendélienne où :
- la dominance est totale,
- un caractère n’est contrôlé que par un seul gène,
- les gènes se transmettent de façon totalement indépendante.
4.1 - Dominance
Cependant, dans la plupart des cas un allèle n’est pas totalement dominant et l’on voit apparaître des intermédiaires.
Dans l’exemple du mono hybridisme, on aura ainsi des feuilles palmées, penniformes et toutes les formes intermédiaires.
4.2 - Interaction génique
De plus, le phénomène dit d’interaction génique existe.
Dans ce cas un caractère est contrôlé par deux ou plusieurs paires de gènes. Les ratios des génotypes sont les mêmes mais les ratios des phénotypes vont varier.
Exemple hypothétique où deux paires d’allèles dominants Pp et Cc contrôlent les anthocyanines (pigments de couleur violet).
Si seul P est présent, seules les feuilles de la plante seront violette. Si seul C est présent, la plante reste verte. Mais si P et C sont présent, les feuilles et les calices de la plante seront violets.
4.3- Transmission indépendante
Quand à la transmission indépendante, elle va dépendre :
- de la position des gènes concernés sur des chromosomes différents ou sur le même chromosome,
- de la distance entre les loci de ces deux gènes si ils sont sur le même chromosome.
Les lois de la génétique mendélienne sont donc assez rapidement dépassées mais offrent une bonne base d’interprétation des résultats des croisements.
5 - Applications a la reproduction sélective
Résumons tout d’abord les règles de base de la sélection :
- les génotypes des plantes sont contrôlés par des gènes qui sont transmis de génération en génération (notion d’hérédité),
- les gènes forment des paires. Dans chaque paire de gène, un allèle est transmis par chacun des parents,
- lorsque les allèles ont un effet différent sur le phénotype, la plante est dite hybride ou hétérozygote,
- quand les allèles d’une paire de gènes exercent la même influence, la plante est dite pure pour la paire de gènes considérée,
- différentes paires de gènes contrôlant différents caractères génotypiques sont (en général) transmises indépendamment,
- les relations de dominance et l’interaction génique peuvent modifier le ratio des phénotypes.
La reproduction sélective consiste en l’hybridation sélective d’individus sélectionnés à l’opposé de l’hybridation au hasard se produisant dans la nature.
Elle a pour but :
- de croiser deux variétés possédant des qualités exceptionnelles,
- de croiser des individus de génération F1 afin de réaliser les potentialités du croisement initial (différenciation),
- rétro croisement (ou backcross) pour rétablir les types parentaux originels,
- croisement de deux variétés vraies (homozygotes) afin de préserver un caractère commun et de restaurer la vigueur de la lignée.
6- La vigueur Hybride
L'hétérosis désigne l'augmentation des capacités et ou de la vigueur d'un hybride par rapport aux races, lignées... dont il est originaire.
L'effet d'hétérosis, nommé également vigueur hybride, se traduit par un gain de performances qui résulte du mélange de différents gènes de différentes races. Le terme est inventé en 1914 et correspond à la découverte du scientifique George Harrison Shull, en élargissant la théorie de son rival Edward East, d'une « stimulation physiologique » due à l'état hétérozygote.
L'effet d'heterosis désigne en génétique l’accroissement particulièrement prononcé de la performance des individus hybrides ou métis. Cet effet est exploité en sélection animale et en sélection végétale.On parlera d’effet d'heterosis lorsque la génération hybride F1 présente des performances supérieures à la performance moyenne de la génération parente P, homozygote ou de lignée pure.
Le phénomène dit de vigueur hybride ou d'hétérosis nous intéresse particulièrement lorsque l'on fait des croisements :
- de deux variétés possédant des qualités exceptionnelles,
- de deux variétés vraies (homozygotes) afin de préserver un caractère commun et de restaurer la vigueur de la lignée.
Si les parents représentent divers patrimoines génétiques, il en résulte une vigueur hybride, parce que les allèles dominants tendent à porter des caractères favorables et que les allèles dominants différents hérités de chacun des parents masquent les caractères récessifs hérités de l'autre.
Evidemment, cela n'est vrai que lorsque la dérive génétique entre les deux lignées parentales est importante.
7 - Hybridation et Différenciation
Dans ce cas, on va croiser des individus de type F1 afin de réaliser une différenciation par le biais de la recombinaison (voir le chapitre dihybridisme).
Le but de la différenciation étant de combiner des qualités différentes provenant des deux parents.
[src]https://www.cannaweed..._8/2/c4b18b.jpg[/src]
{kind=link}
L'hybridation initiale donne un phénotype uniforme dans la génération F1 mais le croisement endogame produit l'apparition de nouveaux phénotypes
8 - Le Cubing
Outre le fait de rétablir les types parentaux originaux, lors d'une perte de vigueur, le rétrocroisement particulier appellé cubing permet d'obtenir à partir d'un seul individu une lignée entière :
- Le premier rétro-croisement est simplement un rétro-croisement.
- Le deuxième rétro-croisement, obtenu en croisant le premier rétro-croisement avec son parent (qui devient donc grand-parent) et souvent appelé SQUARING par les breeders.
- Votre troisième rétro-croisement, obtenu en croisant le deuxième rétro-croisement avec son grand-parent (qui devient donc arrière-grand-parent) et souvent appelé CUBING par les hybrideurs. Vous pouvez continuer le rétro-croisement mais cela s'appelle juste rétro-croisement. Le terme cubing est une référence au chiffre 3, parce qu'il y a 3 rétro-croisements.
Le cubing se base sur des probabilités mathématiques, compte tenu des fréquences des gènes. Plus vous utiliserez de mâles dans vos croisements, plus vous aurez de chances que la réalité colle à la théorie.
En théorie, avec le premier croisement, 75% du pool génétique obtenu correspondra au pool génétique du parent P1. Le squaring monte cette probabilité à 87,5% et le cubing à 93,75%. Vous trouvez ces résultats en faisant la moyenne entre les deux parents croisés.
Par exemple, vous commencez à croiser la mère P1 (100%) avec un mâle quelconque (0%). 100% + 0% divisé par 2 donne 50%. La progéniture sera donc approximativement 50% identique à la mère. Prenez-là, faites votre premier rétro-croisement, et vous obtiendrez 100% + 50% / 2 = 75%.
Et ainsi de suite. Comme nous le verrons plus tard, vous pouvez appliquer ces mêmes probabilités mathématiques à des traits ou des gènes spécifiques, et cela peut avoir un effet dramatique sur votre méthodologie et vos méthodes de sélection.
Votre sélection de mâles pour chaque rétro-croisement est un point crucial pour réussir avec cette technique.
Selon le cas, vous pouvez choisir des mâles qui contiennent les gènes que vous désirez, ou vous pouvez par inadvertance choisir des individus porteurs du gène récessif non-désiré.
Ou plus probablement, vous pouvez choisir des individus hétérozygotes pour les deux gènes comme la mère P1 qui a été rétro-croisée.
La façon de faire la plus facile est de ne s'intéresser qu'à un seul gène et un seul trait, par exemple supposons que le goût est déterminé par un seul gène (ce qui n'est pas le cas en réalité).
Supposons maintenant que parmi notre population qui a un goût de pin nous trouvions un individu spécial qui a un goût d'ananas et que nous voulions le garder. Le gène causant le goût d'ananas peut être dominant ou récessif et les résultats peuvent être différents selon le cas.
8.1 - Le goût d'ananas est dominant
A = goût d'ananas et p = goût de pin
Chaque individu aura deux gènes de goût par paire, les génotypes possibles étant AA, Ap, et pp.
Comme A est dominant, AA et ap exprimeront le goût d'ananas alors que pp donnera le goût de pin, ce sont les phénotypes. Comme l'ananas est un nouveau goût, il y a de fortes chances pour que l'individu spécial soit hétérozygote, à savoir Ap.
Du coup, la seule combinaison de parent possible est Ap X pp, Ap étant le parent que l'on souhaite cuber.
Le croisement F1 :

Il faut maintenant choisir des mâles qui ont un goût d'ananas. Les mâles seront choisis aléatoirement.. Le ratio de A par rapport aux p des mâles F1 sera 2:6 (2 A pour 6 p).
On peut aussi dire que la fréquence du gène A est de 25%. Cela signifie que sur quatre grains de pollen, un contiendra le gène du goût d'ananas.
Voilà ce qui se passe lors du premier rétro-croisement :
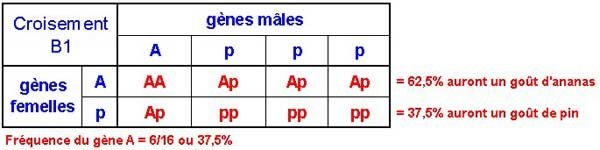
C'est ce premier rétro-croisement qui le premier crée un individu homozygote (AA) pour le goût ananas.
Cependant, à cause de nos moyens de sélection limités, nous choisissons les mâles au hasard. De chaque mâle nous pouvons espérer que trois grains de pollen sur huit contiennent le gène du goût d'ananas.
La femelle P1 continue de donner un A pour un p. Je vais épargner la mémoire de votre ordinateur en ne mettant pas le tableau, mais n'hésitez pas à le faire sur papier pour être sûr d'avoir bien compris ce qu'il se passe.
Le second backcross (Squaring) donnera ceci :
3AA 8Ap 5pp
[p]68,75% auront donc le goût d'ananas et 31,25% le goût de pin. La fréquence du gène A est passé à 7/16 soit 43,75%.
Et finalement, le troisième backcross (le Cubing) donne :
7AA 16Ap 9pp
71,875% ont le goût d'ananas après le cubing. Environ 22% (7 / 32 X 100) de la progéniture est homozygote sur le goût d'ananas. La fréquence du gène A atteint environ 47% (30/64).
En conclusion, si le rétro-croisement continue indéfiniment avec une sélection aléatoire des mâles sur une population suffisamment grande, la fréquence du gène A atteindra au maximum 50%.
Cela signifie que le mieux que nous puissions espérer est 25% de pure souche de goût d'ananas et en tout 75% de plantes qui auront le goût d'ananas. Vous ne vous débarrasserez pas des 25% au goût de pin.
8.2 - Le goût d'ananas est récessif
Dans ce cas, P est le goût de pin et a est le goût d'ananas. La convention est que les lettres majuscules représentent la dominance. La plante sur laquelle on a trouvé le goût d'ananas est forcément homozygote (aa).
Le mâle peut être Pa ou PP, mais plus probablement PP. De toutes façons, cela ne fait pas une grande différence sur les résultats.
Le croisement F1 est plutôt basique, nous nous passerons donc du diagramme. Nous croisons simplement une femelle (aa) avec un mâle (PP) et nous obtenons des individus qui sont tous Pa. Comme le goût d'ananas est récessif, aucun individu de la génération F1 n'aura le goût d'ananas. La fréquence du gène a est cependant de 50%.
aa X PP = Pa + Pa + Pa + Pa
Comme tous les individus F1 sont identiques (Pa), le pollen qu'ils donnent pour le premier rétro-croisement contiendra un gène a pour chaque gène P. Le premier backcross donnera :
B1 = aa X Pa = Pa + Pa + aa + aa[/p]
Comme vous pouvez le voir, 50% de la progéniture a le goût d'ananas et la fréquence du gène a est de 6/8 soit 75%. La génération B1 donne du pollen contenant 6 gènes a pour 2 gènes P.
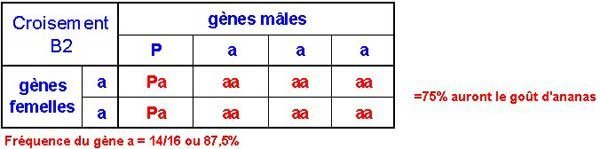
Comme vous pouvez le voir le deuxième rétro-croisement (squaring) donne le goût d'ananas à 75% de la progéniture. La fréquence du gène a est environ 88%.
Sur les grains de pollen, 14 sur 16 porteront le gène a. Quand on les rétro-croise avec la mère P1, on obtient cela :
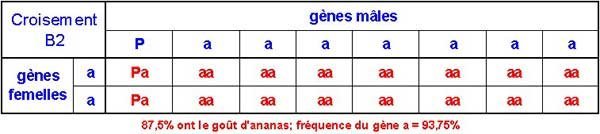
88% environ ont maintenant le goût d'ananas, et toutes sont homozygotes pour ce trait. La fréquence du gène désiré est à peu près de 94%. Si le rétro-croisement continue indéfiniment, la fréquence du gène tendra vers 100%, sans toutefois jamais les atteindre.
Il faut noter que les exemples ci-dessus supposent que l'on n'applique aucun critère de sélection ainsi qu'une population suffisamment vaste pour assurer un choix aléatoire des mâles.
Plus il y a de mâles, meilleure est la sélection, qu'on la veuille aléatoire ou non. L'importance de la taille de la population et des critères de sélection est encore plus grande quand les gènes que l'on souhaite reproduire sont hétérozygotes. Et surtout, nous n'avons pris pour ces exemples qu'une seule paire de gènes.
En réalité, la plupart des traits recherchés comme la puissance sont influencés par plusieurs traits. Les mathématiques deviennent alors plus compliquées pour trouver le taux de succès d'un projet de cubing. En gros, vous devez multiplier les probabilités d'obtenir un trait par les probabilités d'en obtenir un autre.
Par exemple, si le trait du goût d'ananas était influencé par deux gènes récessifs différents, vous devriez alors multiplier 87,5% par 87,5% (0,875X0,875X100) ce qui donne 76,6%. Cela signifie que 76,6% de la progéniture aura le goût d'ananas. Supposons maintenant que le trait du goût d'ananas est influencé par 2 gènes récessifs et un gène hétérozygote dominant. Il faut multiplier 87,5% par 87,5% puis par 71,9% (0,875 X 0,875 X 0,719 X 100) ce qui donne 55%.
En passant juste à trois gènes, le nombre d'individus après le cubing ayant le goût d'ananas tombe à 55%. Le cubing est donc une bonne technique quand vous souhaitez augmenter la fréquence de quelques gènes (il est important de s'en souvenir), mais plus le projet grossit, plus les chances de succès sont faibles… du moins tant qu'on n'applique aucun critère de sélection.
8.3- Appliquer des critères de sélection
Le meilleur moyen d'augmenter significativement vos chances de réussite est d'appliquer des critères de sélection.
Essayez de trouver une façon efficace et précise d'isoler les mâles qui ont les traits désirés et de rejeter ceux qui possèdent des traits non-souhaités. Trouver des moyens d'être sûr que vos mâles transmettent les traits désirés et écartez ceux qui ne le font pas.
TOUS les traits doivent être pris en compte. Certains sont directement observables sur les mâles. D'autres, comme le temps de floraison, non.
Si vous cherchez un trait que vous ne pouvez pas voir directement, vous devez effectuer des tests de progéniture pour déterminer quels mâles transmettent le plus de gènes désirés. Un test de progéniture consiste à mettre en croissance certains individus de la progéniture pour déterminer le génotype parental.
Il est important quand vous choisissez vos meilleurs mâles d'ignorer les traits superficiels qui n'ont rien à voir avec les traits que vous cherchez réellement.
En effet, le cannabis possède plusieurs milliers de gènes sur seulement 10 paires de chromosomes (ou 20 chromosomes individuels). C'est-à-dire que chaque chromosome comporte plusieurs centaines de gènes. Tous les gènes se trouvant sur le même chromosome sont liés entre eux.
En gros, ils voyagent en groupe. Quand vous en choisissez un, vous prenez en fait tous ceux sur le chromosome. Il y a une exception à cette règle qui consiste à casser les liens entre les gènes par croisement, mais au nom de la simplicité, nous n'en parlerons pas. Revenons donc à la sélection.
Vous pourriez choisir un mâle selon un trait que vous aimez bien comme la forme des feuilles en étoile alors que ce que vous cherchez réellement est le goût de pamplemousse.
Mais il se peut que les deux traits se trouvent sur la même paire de chromosomes, mais sur les chromosomes opposés.
Si cela arrive, aussi longtemps que vous choisirez des plantes aux feuilles en étoile, vous n'obtiendrez jamais le goût de pamplemousse que vous désirez réellement.
Il est bien de garder à l'esprit qu'à chaque fois que vous sélectionnez selon un trait, vous en écartez plusieurs centaines.
C'est pourquoi les hybrideurs les plus sérieux apprennent à avancer petit à petit en ne travaillant que sur un trait ou deux à la fois.
8.4 - Cubing sélectif
Maintenant voyons comment nous pouvons améliorer notre premier exemple de cubing d'un trait hétérozygote dominant en appliquant des critères de sélection. Supposons qu'à chaque génération nous sommes capables de retirer les individus homozygotes sur le goût de pin (pp), mais pas les hétérozygotes (Ap).
Si vous vous souvenez, notre mère P1 a le génotype Ap et les combinaisons possibles pour la génération F1 sont Ap + Ap + pp + pp. Nous retirons les deux pp, ce qui ne laisse que les Ap. Notre premier rétro-croisement donnera:
Ap X Ap = AA + Ap + Ap + pp
[p]Encore une fois nous enlevons les pp ce qui nous laisse avec AA + 2Ap. Nous attaquons le deuxième rétro-croisement en ayant passé la fréquence du gène A de 37,5% à 66,7%, 4 grains de pollen sur 6 portent le gène A.
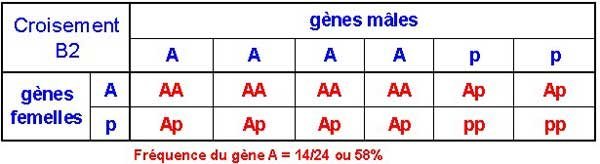
Comme vous pouvez le voir, après avoir éliminé les individus homozygotes récessifs après deux rétro-croisements, la fréquence du gène A est passée de 44% à 58%. Si nous continuons, la fréquence monte à 70% (14/20) avant le troisième rétro-croisement, c'est-à-dire que 7 grains de pollen sur 10 seront porteurs du gène A.
Encore une fois, j'épargnerai la mémoire de votre PC en me contentant de vous donner les résultats du troisième rétro-croisement : B3 = 7AA + 10Ap + 3pp
95% de la progéniture a maintenant le goût d'ananas comparé aux 72% sans appliquer de sélection. La fréquence des individus pure souche goût d'ananas monte à 35%.
Mais plus important, la fréquence du gène A atteint 60%. C'est un point important au moment des tests de progénitures.
8.5 - En conclusion
Récapitulons maintenant les pourcentages d'individus pure souche goût d'ananas obtenus dans chacun de nos modèles.
Dans le cas où le goût d'ananas serait un trait dominant hétérozygote et qu'aucun critère de sélection n'est appliqué, le cubing produit 22% d'individus pure souche.
En éliminant les individus homozygotes récessifs pour le goût de pin, on monte à 35%. Et enfin, quand on cube un gène récessif homozygote, on atteint après le cubing 87,5% d'individus au goût d'ananas pure souche. Et comme je l'ai déjà dit, ces nombres ne sont valables que pour un seul gène.
Supposons que le goût d'ananas soit codé par deux gènes différents, un dominant et un récessif, et que vous soyez capable d'éliminer les individus pp et de ne sélectionner que les individus porteurs du gène dominant du goût d'ananas.
Votre population après cubing contiendra 87,5% X 35% (0,875 X 0,35 X 100) = 30% d'individus pure souche.
Comme vous pouvez le voir, tant que la source est hétérozygote, peu importe le nombre de rétro-croisements que vous effectuez, vous n'obtiendrez jamais une variété pure souche (stabilisée).
9 - La stabilisation
Une fois que l'on a obtenu des individus correspondants à notre recherche, il va falloir stabiliser la lignée. La stabilisation consiste en la fixation des caractères.
Elle se produit soit par le biais du GA3 soit par celui de croisements successifs.
9.1 - Stabilisation à partir d'un seul individu
Il s'agit de stabiliser par le biais d'une autopollinisation. On soumet un individu femelle à un traitement au GA3 ce qui permet d'obtenir des fleurs staminées. Puis on procède à une autopollinisation...
On va donc obtenir des graines exclusivement femelles dites selfed possédant le patrimoine parental pour une très grande partie.
Il va cependant exister une certaine hétérogénéité du fait du brassage intra et inter chromosomique.
9.2 - La stabilisation par croisements successifs
On va procéder aux croisements successifs en gardant les parents potentiels qui correspondent le mieux à nos critères. Il faut au minimum 5 générations avant d'obtenir une lignée à peu près stabilisée.
Par contre, il peut y avoir une perte de vigueur à un moment ou un autre. Il faut donc prendre la précaution de toujours garder les parents des générations précédentes afin de faire un backcross et restaurer les qualités recherchées.
CitationRappelez-vous que dans la production de toute plante, la sélection joue une part décisive. Il faut tout d'abord savoir précisément quel type de plante on souhaite obtenir, puis reproduire et sélectionner à cette fin, en sélectionnant au fil des ans, les plantes qui se rapprochent le plus du type idéal, et en rejetant toutes les autres [....]
L'hérédité est fixée de façon indélébile par la répétition.
10 - Critères de sélection
Après cette brève introduction théorique au breeding, il faut maintenant choisir les caractères sur lesquels vont se baser notre sélection.
3 - Liste des caractéristiques favorables observées chez le cannabis et sujettes à variations
Caractéristiques générales :
- taille et rendement
- vigueur
- adaptabilité
- robustesse
- résistance aux maladies et aux nuisibles
- maturation
- production de racines
- ramification
- sexe
Caractéristiques des jeunes plants
Caractéristiques des feuilles
Caractéristiques des fibres
Caractéristiques florales :
- aspect
- forme
- taille du calice
- couleur
- teneur en cannabinoïdes
- goût et arôme
- persistance des principes aromatiques et des cannabinoïdes
- types de trichomes
- qualité et quantité de résine
- teneur en résine
- séchage et conservation
- facilité de manucure
- caractéristiques des graines
- maturation
- floraison
- mûrissement
- profil cannabinoïde
Phénotypes généraux des variétés de cannabis
11 - La collecte des données
Avant de commencer à mettre en pratique, il va falloir prévoir une méthode de suivi.
A titre d'exemple, le croisement de dix variétés pures pour un seul caractère va donner 10 individus purs et 90 hybrides à la première génération.
Alors, imaginez faire cela sur plusieurs générations et plusieurs caractères.....
Il est donc illusoire de s'en remettre à sa mémoire et il vaut mieux confier le suivi des croisements et leurs résultats au niveau des critères recherchés à un cahier de bord.
12 - La pollinisation
La première étape pratique une fois les parents sélectionnés et les objectifs définis va être la récolte du pollen.
12.1 - Sélection des parents
Il faudra sélectionner les parents de la façon la plus rigoureuse qui soit (cf critères de sélection) et éliminer dès les premières étapes de la croissance végétative les individus les plus faibles et les moins vigoureux.
Il faut aussi essayer d'éliminer les facteurs de monoécie et sélectionner de façon rigoureuse les mâles non seulement sur leur vigueur mais aussi sur leur high et leur rendement en procédant à une inversion sexuelle par le GA3.
12.2 - Récolte du pollen
Attention, le pollen est extrêmement volatile et peut se disperser sur plusieurs kilomètres par le biais du vent, des vêtements ou de vos mains. De plus, la durée de vie des fleurs est brève.
Une méthode alternative consiste à couper les grappes de fleurs mâles avant qu'elles ne s'ouvrent. Vous préparez un récipient avec de l'eau du type utilisé pour vos boutures, recouvrez le d'un film plastique (type cellophane).
Ensuite, percez de petits trous et placez vos grappes de fleurs mâles. Quand elles se sont ouvertes, tapotez et le pollen se répand à la surface et y adhère par électricité statique.
Il ne vous reste plus qu'à le racler à l'aide d'un scalpel. Il est très important de toujours utiliser des instruments stériles.
12.3 - Conservation du pollen
Le pollen ne se conserve que quelques jours à température ambiante. Il faut donc être capable de minuter les passages en floraison de façon raisonnée.
On peut conserver le pollen pour une plus longue durée.
Il faut placer le pollen récolté dans un récipient en verre (type tube sous vide de prise de sang) et y placer un dessicant (type dessicant pour matériell photographique).
Ensuite, on place le pollen deux jours dans un réfrigérateur et enfin on le place dans un congélateur.
Si cette opération est bien faite, on peut augmenter la durée de conservation du pollen jusqu’à quelques semaines.
12.4 - Pollinisation
Il suffit ensuite de le mettre sur le plant femelle sélectionné.
Celui-ci est prêt à être fertilisé lorsque la cosse s’ouvre légèrement pour libérer pistil et styles. Une fois la pollinisation effectuée, une fleur femelle sur deux va avorter.
Les méthodes sont nombreuses, on peut citer le pinceau, ou un sac de toile à mailles très serrées. Si l'on utilise la méthode des sacs, il est conseillé de les enlever au bout de 3 jours puis de les détruire afin d'éviter une pollinisation accidentelle.
13 - Les graines
13.1 - Maturité des graines
Une fois la pollinisation effectuée, il va falloir attendre que la graine se forme. Elle sera mature en quelques semaines et lorsque l'on secoue la grappe florale, elle se séparera d'elle-même.
13.2 - Sélection des graines
Il faut sélectionner les graines :
- selon leur lustre
- leur régularité
- leur robe
- leur taille
- leur lignage, il ne faut sélectionner que les graines dont les parents sont connus et pour lesquelles une pollinisation accidentelle est exclue
- etc....
13.3 - La levée de dormance
Il s'agit de lever la dormance qui est un phénomène de protection des végétaux.
CitationPlacez les graines au noir et au sec pendant 5 jours.
Ensuite placez-les dans le frigo pendant 3 jours.
Ensuite au congélateur pendant 3 jours.
Et de nouveau au noir et au sec pendant 5 jours.
Ensuite, vous n'avez plus qu'à entamer l'étape suivante de votre sélection ainsi qu'à faire germer la prochaine génération.
14 - Conclusion
14.1 - De la théorie à la praqqtique
On n'a abordé ici que les bases élémentaires nécessaires à la compréhension des croisements. La réalité est bien plus complexe que cela.
Et avant de se lancer dans le breeding, une bonne expérience de cultivateur est nécessaire (ne serait-ce que pour arriver jusqu'au stade de la récolte ). De plus, un bon feeling et un peu de chance ne sont pas des facteurs à négliger...
14.2 - Sur combien de pieds travailler ?
Plus les caractères sur lesquels on travaille sont nombreux, plus le nombre de pieds pour chaque génération devra être important. De plus, il est nécessaire de conserver les parents de chaque génération afin de procéder à un back-cross en cas de nécessité.
Et il est parfaitement illusoire de ne vouloir travailler que sur 10 plants....
15 - Le cas particulier de la polyploïdie
Un plant de cannabis polyploïde possède un nombre >2n de chromosomes. En général, ce seront des plantes tétraploïde 4n chromosomes.
15.1 - Conséquence de la polyploïdie
Les travaux de Warmke et al. (1942-1944) montrent : Que les plantes tétraploïdes ont un potentiel double de celui des plantes diploïdes (à tuer des organismes aquatiques).
Mais il n'a jamais mesuré le taux des différents cannabinoïdes qui étaient inconnus à l'époque. une augmentation des phénomènes de monoécie/hermaphrodisme lié aux intersexes XXXX, XYYY, XXXY, XXYY.
15.2 - Obtenir des plants diploïdes
Essayer d'utiliser de la colchicine (inhibiteur de la ségrégation chromosomique dans les cellules filles) est très dangereux car le taux de viabilité lorsque les graines sont traités à concentration efficace est de 4 pour 1000 (zhatov, 1979) et parmi ces graines viables, la plupart seront des plants mosaïques (cohabitation de cellules de nombre chromosomique différent 2n, 3n, 4n etc...) et très peu de véritables tétraploïdes.Sans parler des problèmes à obtenir une population n+1....
Sources :
OverGrow, RIP
Fiches techniques de Canna-Tech (merci Gen&Tik pour ce taff)
La botanique du cannabis de Robert.C CLARKE
Wikipédia
Dad-'s label
-
 5
5
-
 3
3