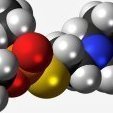
DeltaX9X 164 Posté(e) octobre 11, 2021 Partager Posté(e) octobre 11, 2021 (modifié) Les Chlorophylles Le nom de chlorophylle a été donné en 1818 par P. J. Pelletier et J. B. Caventou aux pigments verts des feuilles. Trente ans plus tard environ, leur parenté chimique avec les pigments sanguins fut soupçonnée, puis la diversité des chlorophylles reconnue. Au début du XXe siècle, commencèrent les travaux qui en révélèrent la structure et permirent d'en réaliser la synthèse en laboratoire. Ces composés organiques liés au magnésium sont engagés dans différents complexes lipoprotéiques membranaires des chloroplastes des cellules végétales. Les Bactéries phototrophes possèdent également des chlorophylles spécifiques (bactériochlorophylles). Ces pigments réalisent les premières étapes de la photosynthèse, c'est-à-dire les étapes photochimiques. Après absorption de photons, les molécules de chlorophylle sont excitées, avec transition électronique, certains de leurs électrons étant expulsés de leur orbite. Les états excités, de courte durée de vie, peuvent donner lieu à une perte d'électrons énergisés, origine d'un transfert oxydo-réducteur. Plusieurs états moléculaires de la chlorophylle assurent ainsi d'une part la collecte énergétique de photons et d'autre part la réduction de transporteurs d'électrons. Cette double fonction permet la transformation du bioxyde de carbone et de l'eau en molécules organiques, c'est-à-dire photosynthèse. Leur rôle de sensibilisateurs de réactions enzymatiques à l'énergie lumineuse fut longtemps traduit par l'expression d'assimilation chlorophyllienne, premier nom de la photosynthèse. A/ Structure et propriétés chimiques : Les chlorophylles sont les pigments verts des végétaux capables de photosynthèse. On en trouve également dans diverses bactéries qui utilisent aussi l'énergie lumineuse. Il en existe 5 sortes de la chlorophylle de la "a" jusqu'à "e". Les chlorophylles les plus communes sont les chlorophylles a et b, présentes dans les chloroplastes des cellules de tous les végétaux de couleur verte : plantes à fleurs, fougères, mousses, algues vertes. Les algues brunes (Fucus, Diatomées) possèdent les chlorophylles a et c, d'autres algues brunes, les Xanthophycées, a et e. Les algues rouges renferment a et d. Ces différentes chlorophylles ne diffèrent entre elles que par de petits détails de structure. Seule la chlorophylle a est constante pour tous les végétaux. Parmi les bactéries phototrophes, l'une, le Prochloron, possède les chlorophylles a et b ; les autres ont des bactériochlorophylles qui leur sont propres. Tous ces pigments ont une structure chimique semblable ; tous comprennent un métal, le magnésium. Les feuilles renferment environ 1 g de chlorophylles pour 100 de substance sèche, soit 1 à 2 pour 1000 de substance fraîche. Elles ont deux fois plus de a que de b. Les chlorophylles sont responsables d'une partie importante, souvent majeure, de l'absorption de la lumière par les végétaux. Pure, la chlorophylle a se présente en aiguilles cristallines bleu sombre, sa formule brute est C55H72O5N4Mg. La chlorophylle b est vert foncé. Les molécules des chlorophylles ont une masse moléculaire voisine de 900. Elles sont toutes formées de quatre noyaux pyrrole I, II, III, IV liés entre eux. Un cycle cyclopentanone (V) est accroché au noyau pyrrole III. Toutes les molécules contiennent encore un groupe dérivé du méthanol et un groupe dérivé d'un alcool à vingt atomes de carbone, le phytol. Ce sont des esters de méthanol et de phytol. Après libération de ces deux alcools par hydrolyse alcaline, il reste l'ensemble tétrapyrrolique lié au magnésium, ou chlorophylline. Une enzyme présente dans les cellules végétales, la chlorophyllase, catalyse le décrochement du seul phytol et laisse une chlorophyllide, étape également de la synthèse des chlorophylles. Enfin, l'action des acides décroche le magnésium et les composés bruns obtenus sont des phéophytines. Les noyaux pyrroliques possèdent de courtes chaînes latérales, vestiges du mécanisme de leur formation. Les chlorophylles b, c, d, e et la bactériochlorophylle ne diffèrent de la chlorophylle a que par la nature de ces courtes chaînes ; ainsi, la chlorophylle b possède un groupe – CHO sur le noyau pyrrole, à la place d'un – CH3. De telles différences en entraînent aussi d'importantes dans les spectres d'absorption de la lumière. Les chlorophylles sont solubles dans les solvants des lipides tels que les alcools, l'éther ordinaire, l'éther de pétrole, l'acétone. La technique de la chromatographie, si employée maintenant dans la séparation et la purification des substances chimiques, a pour origine la séparation des chlorophylles a et b des feuilles. Elle fut réalisée en 1906 par le botaniste M. S. Tswett ; après avoir versé une solution des pigments des feuilles sur une colonne de carbonate, il constata que les chlorophylles et les caroténoïdes se séparaient les uns des autres, étant entraînés par le solvant à des vitesses différentes. Molécule Complexe chlorophyllienne. . Les chlorophylles se décolorent par oxydation. Avec les sels ferriques, l'oxydation est réversible et les oxychlorophylles formées peuvent être réduites et régénérer les chlorophylles. Mais les oxydants énergiques provoquent une décoloration irréversible. Il en est de même de la lumière intense, agent d'une photo-oxydation qui peut aboutir à la décoloration complète des feuilles (solarisation). On a attribué aussi aux chlorophylles des propriétés désodorisantes dues sans doute à ce qu'elles fixent différentes substances et les soustraient à l'odorat. Il faut également souligner la parenté chimique entre la chlorophylle, l'hémochromogène (ou hème de l'hémoglobine) et l'hématine qui en dérive. Il s'agit toujours de molécules à quatre noyaux pyrroliques. Mais dans l'hème, ces noyaux sont liés à un atome de fer et il n'y a pas de groupes phytol et méthanol. Les pigments bleus et rouges des algues bleues et des algues rouges sont des protéines, les phycocyanines et phycoérythrines. Ils doivent leur couleur à des composés, ou chromophores, également tétrapyrroliques. Les noyaux pyrrole y sont unis entre eux, mais ne forment pas un ensemble fermé sur lui-même. Ils ne sont pas liés à un métal. Par leur structure, ces chromophores sont très voisins des pigments biliaires. B/ Propriétés Electromagnétiques des chlorophylles : La richesse des molécules de chlorophylles en doubles liaisons conjuguées entre les atomes de carbone et d'azote, doubles liaisons séparées par une seule liaison simple ( = C − C = ou = C − N = ), leur communique une intense coloration. En solution dans l'éther, l'acétone ou le méthanol, la chlorophylle a est bleu-vert alors que la chlorophylle b est vert-jaune. Ces pigments possèdent deux bandes d'absorption intense. *** L' une concerne les radiations bleues (420-480 nm ), elle est commune à tous les pigments tétrapyrroliques (bande de Soret). *** l'autre se situe dans la partie rouge du spectre (640-680 nm). Leurs spectres montrent l'importance de l'absorption des radiations visibles dans le bleu et le rouge. Dans les chloroplastes où les chlorophylles sont concentrées en agrégats, on observe un décalage des maximums d'absorption vers les grandes longueurs d'onde. L'agrégation des molécules de chlorophylles entre elles et avec d'autres molécules, en complexes colloïdaux, s'accompagne en effet d'un déplacement des maximums d'absorption de 10 à 15 nanomètres vers les grandes longueurs d'onde. Les solutions colloïdales aqueuses de chlorophylles présentent un déplacement du même ordre. Une partie de la diffusion de la lumière par les cellules végétales est due à un tel état d'agrégation. Quant aux bactériochlorophylles, leurs maximums d'absorption sont situés dans le proche infrarouge. In vivo, ils se situent à 800, 850, 890 nm, pour les bactéries pourpres. Les solutions de chlorophylles présentent une belle fluorescence rouge correspondant à une émission de radiations de plus grande longueur d'onde que celle de la lumière absorbée. Cette fluorescence est diminuée par l'addition de quinone qui joue le rôle d'extincteur (quencher). Les monocouches de molécules de chlorophylles juxtaposées, obtenues à partir de solutions inertes convenablement étalées et dont le solvant a été évaporé, ne sont pas fluorescentes tandis que les chlorophylles in vivo le sont. On ne peut donc assimiler leur état biologique à celui de monocouches amorphes. C/ Synthèse, biogenèse : La synthèse de la chlorophylle a a été complètement réalisée in vitro en 1960 par R. B. Woodward et son équipe à Harvard, en plusieurs étapes. Le principe de la méthode comprend la synthèse des noyaux pyrrole, leur accrochage entre eux, leur réduction partielle, l'addition progressive des chaînes latérales, des plus courtes aux plus longues. Mais les moyens empruntés à la chimie organique de synthèse sont très différents du mécanisme de la biogenèse. Ce dernier, pour la synthèse du pyrrole, utilise des métabolites banaux : l'acide succinique, composé intermédiaire du métabolisme respiratoire, et un acide aminé, le glycocolle. Leur condensation en acide δ-aminolévulinique et la condensation de deux molécules de ce dernier donne un noyau pyrrole déjà muni de ses chaînes latérales, ou porphobilinogène. D/ Excitation photochimique des chlorophylles et photosynthèse : L'excitation des molécules de chlorophylle par les photons peut provoquer plusieurs phénomènes selon la stabilité des électrons concernés et la longueur d'onde de la lumière absorbée. Seuls les électrons π, faiblement liés au squelette moléculaire, sont excités par les radiations visibles. Différents états ou niveaux énergétiques de la chlorophylle, ou excitons, ont pu être caractérisés, soit dans les organismes eux-mêmes, soit dans des cristaux de chlorophylle, par l'étude des spectres à basse température, de la polarisation, de la fluorescence, du spectre de diffusion Raman de résonance des différentes formes pigmentaires. Leur formation et leurs transformations sont extrêmement rapides. Ainsi un électron peut être expulsé de son orbite, la molécule ayant subi une transition entre son état électronique fondamental stable et un état excité. Pour la chlorophylle, il existe deux niveaux énergétiques correspondant à l'absorption de photons dans le bleu-violet, d'une part, et deux correspondant à l'absorption dans le rouge, d'autre part. Or un électron appartient toujours à une paire ; les moments angulaires de spin des deux électrons d'une paire peuvent rester antiparallèles, ce qui définit l'état singulet.La durée de vie d'une molécule excitée par des photons violets qui, dans le visible, permettent d'atteindre l'état énergétique le plus élevé, est inférieure à 5 . 10-12 seconde. Dans le cas d'excitation par la lumière rouge, moins énergétique, l'état atteint a une durée de vie plus longue, de 1 à 5 . 10-9 seconde. La molécule à l'état singulet le plus élevé peut se convertir en l'état le plus bas en 10-15 seconde, avec émission de chaleur : C*v → C*r + chaleur. L'état énergétique le plus bas C*r peut retourner à l'état fondamental, avec émission de lumière rouge, origine de la fluorescence rouge de la chlorophylle in vivo et in vitro, et de chaleur. Les états excités peuvent aussi transférer leur énergie à des molécules voisines, soit par collision, soit par résonance. Ces transferts ne sont efficaces que si les molécules sont très rapprochées les unes des autres, comme c'est le cas in vivo où les molécules de chlorophylle sont concentrées dans les membranes des chloroplastes. Enfin, ces états excités singulets peuvent se transformer en un autre état énergétique, l'état triplet, pour lequel les moments angulaires de spin sont parallèles. L'état triplet a une durée de vie beaucoup plus longue, de l'ordre de 1 . 10-3 seconde. On le dit métastable. À partir de cet état, le retour à l'état fondamental peut s'effectuer avec dégagement de chaleur ou par transfert d'énergie à une autre molécule pigmentée. Un autre type de transformation possible a été également mis en évidence. Il s'agit d'une oxydo-réduction dans laquelle intervient une séparation de charge, un électron quittant la molécule de chlorophylle qui se trouve alors ionisée, chargée positivement. Elle peut ainsi intervenir dans une série de transferts d'électrons entre un donneur, l'acide ascorbique par exemple, et un accepteur, une quinone, selon la séquence : Chaque transition électronique est caractérisée par son énergie E, fonction de la fréquence ν de la radiation d'excitation (E = hν ; h : constante de Planck, et ν = c/λ ; c : vitesse de la lumière ; λ : longueur d'onde). Le passage de la molécule de son état stable à l'état excité C* est réalisé dans un temps très court, correspondant à la période de vibration de la lumière, soit de 10-14 à 10-15 seconde. Ainsi, la réduction de la quinone par l'ascorbate, qui n'aurait pas lieu spontanément en raison de son besoin énergétique, se trouve-t-elle sensibilisée à l'énergie lumineuse par la chlorophylle excitée, « pont » électronique entre l'énergie électromagnétique de la lumière et l'énergie chimique. D'autres donneurs d'électrons et d'autres accepteurs peuvent être ainsi mis en relation et ce modèle simple constitue une image schématique du processus d'oxydo-réduction des transporteurs d'électrons par l'eau, au cours de la photosynthèse. In vivo, un dimère de chlorophylle, dont le maximum d'absorption dans la lumière rouge est à 700 nm et appelé pour cette raison P700, est ainsi capable, après éclairement, d'assurer la réduction d'un transporteur biologique, le nicotinamide-dinucléotide phosphate qui se charge d'électrons + protons. Ce dimère ne représente qu'environ un pour cent des molécules totales de chlorophylle présentes. Un autre dimère de chlorophylle, P682, intervient également dans les transferts d'électrons qui prennent leur origine dans les ions OH- venant de la dissociation de l'eau. Ces deux dimères de chlorophylle appartiennent aux centres actifs, fonctionnels, des deux photosystèmes de la photosynthèse. À côté de ces centres actifs, et pouvant en être séparés par voie d'analyse chimique, par électrophorèse, en raison de leur couplage avec des protéines, la majeure partie des molécules de chlorophylle a ainsi que la chlorophylle b chez les végétaux verts, une partie des caroténoïdes, dont le fucoxanthol des algues brunes, et enfin les pigments rouges (phycoérythrines) et bleus (phycocyanines) des algues rouges et des cyanobactéries constituent une antenne collectrice de photons. Dans cette antenne, les transferts d'excitons par résonance apparaissent de règle. Ces transferts sont démontrés par la possibilité de provoquer la fluorescence de la chlorophylle a, dans un mélange des pigments cités, avec de la lumière absorbée d'une manière exclusive par ces autres pigments. Les rendements de transfert sont de l'ordre de 90 %. La pluralité pigmentaire étend la gamme des longueurs d'onde de la lumière pouvant être absorbée. Les transferts en canalisent l'énergie vers les centres actifs où débute le cheminement des transferts d'électrons générateurs de la réduction du bioxyde de carbone en molécules organiques.La photosynthèse se caractérise matériellement par un transfert d'électrons et de protons de l'eau au dioxyde de carbone qui se trouve réduit avec formation de glucides. Cette opération requiert de l'énergie et, dans les meilleures conditions, il faut 8 photons-grammes (8 einsteins) par molécule-gramme de bioxyde de carbone réduit, soit l'équivalent d'environ 1 460 à 2 300 kilojoules, selon la longueur d'onde de la lumière. Le passage de la molécule de son état stable à l'état excité C* est réalisé dans un temps très court, correspondant à la période de vibration de la lumière, soit de 10-14 à 10-15 seconde. Or un électron appartient toujours à une paire ; les moments angulaires de spin des deux électrons d'une paire peuvent rester antiparallèles, ce qui définit l'état singulet. La durée de vie d'une molécule excitée par des photons violets qui, dans le visible, permettent d'atteindre l'état énergétique le plus élevé, est inférieure à 5 . 10-12 seconde. Dans le cas d'excitation par la lumière rouge, moins énergétique, l'état atteint a une durée de vie plus longue, de 1 à 5 . 10-9 seconde. La molécule à l'état singulet le plus élevé peut se convertir en l'état le plus bas en 10-15 seconde, avec émission de chaleur : C*v → C*r + chaleur. L'état énergétique le plus bas C*r peut retourner à l'état fondamental, avec émission de lumière rouge, origine de la fluorescence rouge de la chlorophylle in vivo et in vitro, et de chaleur. E/ Memento schématique : Modifié octobre 11, 2021 par DeltaX9X Lien à poster Partager sur d’autres sites
DeltaX9X 164 Posté(e) mars 28, 2022 Auteur Partager Posté(e) mars 28, 2022 X Lien à poster Partager sur d’autres sites

Messages recommandés